The basic type of plastid in the algae is a chloroplast, a plastid capable of photosynthesis. Chromoplast is synonymous with chloroplast; in the older literature a chloro plast that has a color other than green is often called a chromoplast. A proplastid is a reduced plastid with few if any thylakoids.
A proplastid will usually develop into a chloroplast although in some heterotrophic algae it remains a proplastid. A leucoplast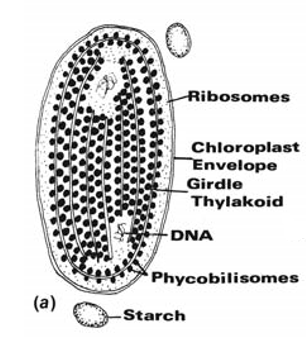 or amyloplast is a colorless plastid that has become adapted for the accumulation of storage product.
or amyloplast is a colorless plastid that has become adapted for the accumulation of storage product.
In the Rhodophyta and Chlorophyta, the chloroplasts are bounded by the double membrane of the chloroplast envelope (Fig a).
In the other eukaryotic algae, the chloroplast envelope is surrounded by one of two membranes of chloroplast endoplasmic reticulum (chloroplast E.R.), which has ribosomes attached to the outer face of the membrane adjacent to the cytoplasm.
The chloroplast E.R. is the remnant of the food vacuole membrane and/or the plasma membrane involved in the original endosymbiosis leading to the chloroplasts in a secondary endosymbiosis.
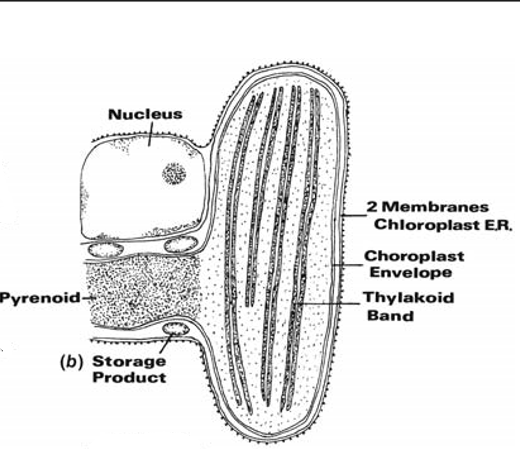
Fig (b) Two thylakoids per band, two membranes of chloroplast E.R. (Cryptophyta).
In the Euglenophyta and Dinophyta, there is one membrane of chloroplast E.R. (Fig. 1.12(c).
In the Cryptophyta, Prymnesiophyta, and Heterokon – tophyta, there are two membranes of chloroplast E.R., with the outer membrane of chloroplast E.R. usually continuous with the outer membrane of the nuclear envelope, especially if the chloroplast number is low.
The basic structure of the photo syn thetic apparatus in a plastid consists of a series of flattened membranous vesicles called thylakoids or discs, and a surrounding matrix or stroma. The thylakoids contain the chlorophylls and are the sites of the photochemical reactions; carbon dioxide fixation occurs in the stroma.
The thylakoids can be free from one another or grouped to form thylakoid bands. In the cyanobacteria and Rhodophyta (Fig. (a)), the thylakoids are usually free from one another, with phycobilisomes (containing the phycobiliproteins) on the surface of the thylakoids. The phycobilisomes on the surface of one thylakoid alternate with those on the surface of an adjacent thylakoid. The phycobilisomes appear as 35-nm granules when phycoerythin predominates, or as discs when phycocyanin predominates.
In the Cryptophyta, the chloroplasts contain bands of two thylakoids (Fig. (b)); the phycobiliproteins are dispersed within the thylakoids.
In the Euglenophyta and Heterokontophyta the thylakoids are grouped in bands of three with a girdle or peripheral band running parallel to the chloro plast envelope.
In the Dinophyta, Prymnesiophyta, and Eustigmatophyceae, the thylak oids are also in bands of three, but there is no girdle band (Fig. 1.12(c), (d)). In the Chlorophyta, the thylakoids occur in bands of two to six, with thylakoids running from one band to the next. The above grouping of algal thylakoids into bands occurs under normal growth conditions. Abnormal growth conditions commonly cause lumping of thylakoids and other variations in structure.
Pyreniods:
A pyrenoid is a differentiated region within the chloroplast that is denser than the surrounding stroma and may or may not be traversed by thylakoids. A pyrenoid is frequently associated with storage product. Pyrenoids contain ribulose-1, 5-bisphosphate carboxylase/oxygenase (Rubisco), the enzyme that fixes carbon dioxide. Consequently, the size of the pyrenoid will vary depending on how much Rubisco is present.
Rubisco exists in two forms
1 Form I occurs in some bacteria, the cyanobacteria, in all green plants and nongreen plants. Form I is composed of eight large subunits and eight small subunits. Form I has a high affinity for CO2 and a low catalytic efficiency (low rate of CO2 fixation). In green algae, euglenoids, and green plants, the large subunit is coded by chloroplast DNA and the small subunit by nuclear DNA. In the cyanelle (endosymbiotic cyanobacterium) of Cyanophora paradoxa and in some non-green algae, both subunits are coded by chloroplast DNA.
2 Form II occurs in some eubacteria and in the dinoflagellates and is composed of two large sub units. Form II has a low affinity for CO2 and a high catalytic efficiency. The common ancestor of all ribulose-1,5-bisphosphate carboxylase was probably similar to Form II and was adapted to the anaerobic conditions and high CO2 concentrations prevailing in the ancient earth. Form I evolved as the earth’s atmosphere became oxygenated, and CO2 concentration declined and with it the need for a greater affinity for CO2. The greater affinity for CO2 in Form I, however, came at the price of reduced catalytic efficiency.
Many motile algae have groups of tightly packed carotenoid lipid-globules that constitute an orange-red eyespot or stigma that is involved in response to light. Motile algae exhibit three types of responses to light