Occurrence:

Chara
Chara is a fresh water, green alga found submerged in shallow water ponds, tanks, lakes and slow running water. C. baltica is found growing is brackish water and C. fragilis is found in hot springs. Chara is found mostly in hard fresh water, rich in organic matter, calcium and deficient in oxygen. Chara plants are often encrusted with calcium carbonate and hence are commonly called stone wort.
Chara often emits disagreeable onion like odour due to presence of sulphur compounds. C. hatei grows trailing on the soil C. nuda and C. grovesii are found on mountains, C. wallichii and C. liydropitys are found in plains.
The fossils of alga have been discovered from Paleozoic era.
Plant Body of Chara:

Fig: 2; Chara Web
The thallus of Chara is branched, multicellular and macroscopic. The thallus is normally 20-30 cm. in height but often may be up to 90 cm to l m. Some species like C. hatei are small and may be 2-3 cm. long.
The plants in appearance resemble Equisetum hence Chara is commonly called as aquatic horsetail. The thallus is mainly differentiated into rhizoids and main axis.
Rhizoids:
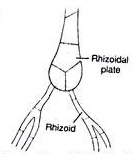
Fig 3: Rhizoids of Chara
The rhizoids are white, thread like, multicellular, uniseriate and branched structures. The rhizoids arise from rhizoidal plates which are formed at the base of main axis or from peripheral cells of lower nodes. The rhizoids are characterized by presence of oblique septa The tips of rhizoids possess minute solid particles which function as statoliths.
The rhizoids show apical growth. Rhizoids help in attachment of plant to substratum i.e., mud or sand, in absorption of minerals and in vegetative multiplication of plants by forming bulbils and secondary protonema.
The main axis:
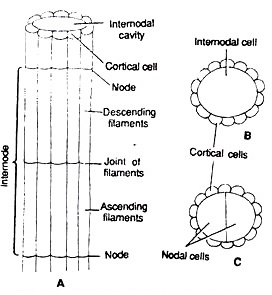 The main axis is erect, long, branched and differentiated into nodes and internodes.
The main axis is erect, long, branched and differentiated into nodes and internodes.
The internode consists of single, much elongated or oblong cell.
The inter-nodal cells in some species may be surrounded by one celled thick layer called cortex and such species are called as corticate species.
The species in which cortical layer is absent are called ecorticate species.
The node consists of a pair of central small cells surrounded by 6-20 peripheral cells. The central cells and peripheral cells arise from a single nodal initial cell.
There are two types of branches in the Chara
(i) Branches of limited growth.
The branches of limited growth arise in whorls of 6-20 from peripheral cells of the nodes of main axis or on branches of unlimited growth. These are also called branchlets, branches of first order, primary laterals or leaves. These branches stop to grow after forming 5-15 nodes and hence are called branches of limited growth. The stipulodes and reproductive structures are formed on the node of these branches.
(ii) Branches of unlimited growth:
The branches – of unlimited growth arise from the axils of the branches of limited growth hence these are also called auxiliary branches or long laterals. These are differentiated into nodes and internodes. At nodes they bear primary laterals and these branches look like the main axis. Their growth is also unlimited like main axis.
(iii) Stipulodes:
The basal node of the branches of limited growth develops short, oval, pointed single cell outgrowths called stipulodes. In most of the species of Chara e.g., C. burmanica, the number of stipulodes at each node is twice the number of primary laterals, such species are called as bi-stipulate.
In some species of Chara e.g., C. nuda and C. braunii, the number of stipulodes at each node, is equal to number of primary laterals at that node, such species are called unitipulate. When stipulodes are present in one whorl at each node the species are called as haplostephanous and with two whorls on each node are called diplostephanous.
Cortex:
If the inter-nodal cells of main axis en-sheathed by cortex cells. Such species are called corticated species.
The cortex consists of vertically elongated narrow cells. The internode up to half of its length by corticating filaments developed from upper node called descending the lower half of internode is covered by filaments developed from lower node called filaments. The ascending and descending filaments meet at the middle of internode. The species without cortex e.g., C. corallina are called ecorticated species.
Cell Structure of Chara:
The main axis of Chara consists of mainly two types of cells:
(i) Nodal cells
(ii) Inter-nodal cells.
The nodal cells are smaller in size and isodiametric. The cells are dense cytoplasmic, uninucleate with few small ellipsoidal chloroplasts. The central vacuole is not developed instead many small vacuoles may be present. The cytoplasm can be differentiated in outer exoplasm and inner endoplasm (Fig. 5 A).

The inter-nodal cells are much elongated. The cytoplasm is present around a large central vacuole. The cells are multinucleate and contain many discoid chloroplasts. The cytoplasm is also differentiated into outer exoplasm and inner endoplasm. The endoplasm shows streaming movements (Fig. 5 B). The cell walls between the nodal cell and inter-nodal cells are porous to help in cytoplasmic continuity between cells.
Reproduction in Chara:
Reproduction in Chara takes place by vegetative and sexual methods. Asexual reproduction is absent.
(i) Vegetative Reproduction in Chara:
Vegetative reproduction in Chara takes place by following methods:
(a) Bulbils:
The bulbils are spherical or oval tube-like structures which develop on rhizoids. C. aspora or on lower nodes of main axis e.g., C. baltica. The bulbils on detachment from plants germinate into new thallus (Fig. 6 A).
(b) Amylum Stars:
In some species of Chara e.g., C. stelligna, on the lower nodes of main axis develop multicellular star shape aggregates of cells (Fig. 6 B). These cells are full of amylum starch and hence are called Amylum stars. The amylum stars do detachment from plants develops into new Chara thalli.

(c) Amorphous bulbils:
The amorphous bulbils are group, many cells, irregular in shape which develop on lower node main axis e.g., C. delicatula or on rhizoids e.g., C. fragifera and C. baltica. The amorphous bulbils are perennating structures, when the main plant dies under unfavorable conditions; these bulbils survive and make Chara plants on return of favourable conditions.
(d) Secondary Protonema:
These are tubular or filamentous structure which develops from primary protonema or the basal cells of the rhizoids. The secondary protonema like primary protonema form Chara plants.
Sexual Reproduction in Chara:
The sexual reproduction in Chara is of highly advanced oogamous type. The sex organs are macroscopic and complex in organization.
The male sex organs are called antheridium or globule and the female oogonium or nucule. Most of the Chara species are homothallic i.e., the male and male sex organs are borne on the same nodes, (Fig. 7) e.g., C. zeylanica. Some species e.g., C. wallichii are heterothallic i.e., male and female sex organs are borne on different plants.
The sex organs arise on the branches of limited growth or primary laterals, the nucule above the globule. The development of globule and nucule takes place simultaneously but species globule matures before nucule (Fig. 7 A, B).

Globule:
The globule is large, spherical, red or yellow structure.
Development and Structure:
The early development of globule and nucule is similar.
The pripheral cell of the lower node of the primary lateral divides periclinally to form and an inner cell.
The outer cell functions as antheridial initial and the lower cell again divides a periclinal division.
Out of these three cells formed,
the lowermost functions as inter-nodal cell
the middle forms basal node,
the uppermost functions as the antehridial initial (Fig. 8 A, B).
The middle basal node cell divides to make 5 peripheral cells. Out of these five peripheral cells, the upper one develops into oogonium, two lateral ones form unicellular bracteoles and two lower ones, one on either side of oogonium forms cortex or remains non-functional (Fig. 8 C, D).

The antheridial initial divides by transverse division to make basal pedicel cell and a terminal antheridial mother cell (Fig. 9 A, B). The pedicel cell does not divide further and forms pedicel or stalk of mature antheridium. The antheridial mother cell enlarges and divides by two successive vertical division at right angle to each other to make a quadrant (Fig. 9 C-D).
All these four cells divide by a transverse wall to make eight cells or octant stage. Each cell of the octant divides periclinally and forms two layers of eight cells each.
The cells of inner or outer layer divide periclinally to make three radial layers of eight cells each. The outermost eight cells enlarge laterally to form a curved plate of eight shield cells (Fig. 9 E, F). The cells of the middle layer elongate towards centre to make eight rod-shaped manubrial cells (Fig. 9 F).
The cells of the inner layer function as eight primary capitulum cells. Each primary capitulum cell divides to form six secondary capitulum cells. Sometimes the secondary capitulum cells divide to make tertiary capitulum cells.
Each capitulum cell divides repeatedly to form 2-4 long, multicellular, branched or un-branched antheridial Filaments or spermatogenous filaments (Fig. 9 G-H). The antheridial filament has up to 250 uninucleate cells. These cells function as sperm mother cell and each cell gives rise to a single spirally coiled, uninucleate, bi-flagellated antherozoid (Fig. 9 J-L).
The mature globule thus is made up of 8 curved shield cells, 8 elongated manubrial cells, 8 centrally located primary capitulum cells and 48 secondary capitulum cells. The secondary capitulum cells give rise to many antheridial filaments. Each sperm mother cell forms a single bi-flagellated antherozoid (Fig. 9 K).

Liberation of Antherozoids:
At maturity the shield cells of antheridium separate from each other exposing antheridial filaments in water. The sperm mother cell gelatinizes to liberate the antherozoids.
Nucule:
The nucule of Chara is large, green, oval structure with short stalk. It is borne at the node of the primary lateral. It lies just above the globule in homothallic species.
Development and Structure:
The upper peripheral cell of the basal node of the antheridium functions as the oogonial initial. The oogonial initial divides by two transverse divisions to make three celled filament. It has lower pedical cell, the middle nodal cell and the upper oogonial mother cell (Fig. 10 A-C).
The pedicell does not divide further and makes pedicel of the oogonium. The middle nodal cell under many vertical divisions to make five sheath cells or peripheral cell which surrounds the central cell (Fig. 10 D-F). The central cell does not divide and functions as the node of the oogonium.

The oogonial mother cell elongates vertically and divides by transverse division to make lower small stalk cell and an upper large oogonium. The oogonium contains uninucleate ovum or egg (Fig. 10 G, H). The peripheral cells or five sheath initials elongate and divide by transverse division to make two tiers of five cells each.
The five upper tier cells from coronary cells which form the corona of nucule. The five lower tier functions as tube cells, the tube cells elongate and get spirally twisted in clockwise directions on oogonium. The nucleus migrates on lower side and receptive spot develops at the tip of oogonium. Large amount of starch and oil get deposited in oogonium.
The mature nucule is attached to the node by the pedicel cell. The nucule is surrounded by five tube cells. The tips of tube cells from corona at the top of nucule. The oogonial cell possesses a single large egg or ovum. The nucule contains large amount of starch and oil. The receptive spot is present at the upper part of nucule.
Fertilization:
When the oogonium is mature, the five tube cells get separated from each other forming narrow slits between them.
Antherozoids are chemotactically attracted towards ovum. The antherozoids enter through these slits and penetrate gelatinized wall of the oogonium. Many antherozoids enter oogonium but one of those fertilizes the egg to make a diploid zygote. The zygote secretes a thick wall around itself to make oospore.
Oospore:
The mature oospore is hard, oval, ellipsoid structure which may be brown e.g., C. inferma, black e.g., C. corallina or golden brown e.g., C. flauda. The oospore inside contains a diploid nucleus and many oil globules in cytoplasm.
On maturity of oospore the inner walls of tube cells get thickened, suberised and silicified. The oogonial as well as oospore walls become thick. The oospore nucleus moves towards the apical region. In advanced stage the outer walls of the envelope or sheath cells fall off and the inner parts remain attached to mature oospore in form of ridges.
Germination of Oospore:
The oospore germinates when favourable conditions appear. The diploid nucleus present in apical colourless region divides by meiosis forming four haploid daughter nuclei (Fig. II A-B).
At this stage a septum divides oospore into two unequal cells. The upper smaller apical cell contains a single nucleus and the large basal cell contains three nuclei. The three nuclei of basal cell degenerate gradually. The oospore apical cell divides by longitudinal division to make a rhizoidal initial and protonemal initial (Fig. 11 D).
The rhizoidal initial shows positive geotropism and forms primary rhizoid, the protonemal initial shows negative geotropism and forms primary protonema. The primary protonema differentiates into nodes and internodes. The peripheral cells of the basal node give rise to rhizoids and secondary protonema. The peripheral cells of the upper nodes give rise to lateral branches (Fig. 11 E-I).

Life Cycle of Chara:
The plant body of Chara is haploid. The vegetative reproduction takes place by the formation of amylum stars, bulbils and secondary protonema. Asexual reproduction is absent. The sexual reproduction is advanced oogamous type.
The male and female sex organs are globule and nucule respectively. After fertilization a diploid spore is formed. At the time of germination diploid oospore nucleus divides to make hapoid nuclei and haploid Chara plant. Thus the life cycle of Chara a predominantly haploid type (Fig. 12, 13).

