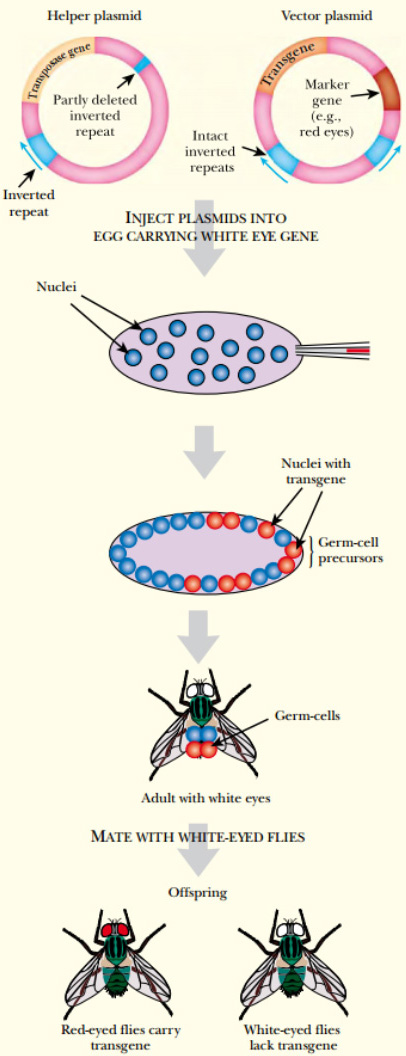
FIGURE P Element Engineering in Drosophila Two different plasmids are used to insert transgenes into Drosophila . The helper supplies the transposase. It carries an immobile P element with one of the inverted repeats deleted but with functional transposase. The second plasmid has the transgene plus marker (a gene for red eyes) flanked by the P element inverted repeats. Both vectors are injected into the posterior end of an egg, which has 2000 to 4000 nuclei within one membrane. The transposase is expressed, which results in random transposition of the transgene (plus marker) into various chromosomes in different nuclei. Hopefully, some insertions occur in germ cell nuclei. The egg is then allowed to form an adult (with white eyes in this case). This fly is then mated to another white-eyed adult. If insertion into the germline was successful, some offspring will express the marker gene and have red eyes.
Several insects can now be genetically modified. The fruit fly Drosophila has been investigated at the molecular level for a long time and, not surprisingly, methods exist for introducing novel genetic material into these flies. P elements are transposons found in Drosophila and other insects, where they cause hybrid dysgenesis . In flies carrying a P element, the frequency of transposition is very low because of synthesis of a repressor protein encoded by the resident P element. When P-carrying males are crossed with P-negative females, the transposition frequency in the fertilized egg is very high for a brief period, due to lack of repressor. Random insertion of P elements then causes a high mutation rate and lowers the proportion of viable offspring, that is, hybrid dysgenesis. P elements are flanked by perfect 31 base-pair inverted repeats. Any DNA sequence that is included between these inverted repeats will be transposed. Therefore engineered P elements can be used to introduce any sequence of DNA into a strain of fruit flies or other susceptible insects. DNA may be microinjected into embryos of P-negative strains of Drosophila . In fruit flies the diploid nucleus resulting from fusion of the sperm and egg nuclei divides multiple times without cell division, resulting in a giant cell with many nuclei, known as a syncytium . Microinjection is normally done at this stage, and incoming DNA usually integrates into at least some of the nuclei that will give rise to the future germline cells. These nuclei are clustered at one end of the fertilized egg—the posterior end . The nuclei then migrate to the outer membrane where a cleavage furrow forms around each nucleus. These furrows expand to form individual cells for each nucleus. The center part remains undivided and acts as a yolk, providing nutrients to the developing larva. The incoming P element is normally carried on a bacterial plasmid that was constructed in a bacterial host. The P element transposes into the Drosophila chromosomes, and the plasmid sequences are left behind. In practice, two P elements are often used. One, the helper, provides the transposase but cannot itself move because of defective 31 base-pair inverted repeats. The other P element, the vector, carries the desired transgene and has intact 31 base-pair inverted repeats, but lacks the transposase gene. Transposition of the vector depends on transposase made by the helper. Once the P element vector has inserted into a particular location on the insect chromosome, it cannot move in future cell generations, because it has no transposase of its own. Ideally, it will be inherited stably. The presence of the P element is monitored by appropriate marker genes. Selectable markers used in flies include neo (neomycin resistance) and adh (alcohol dehydrogenase). Alternatively, eye color genes may be used to reveal the presence of a P element vector. Eye color cannot be positively selected; instead, flies are screened for changes in eye color. For example, flies defective in the rosy gene may be used as host. These flies have brown eyes because of lack of xanthine dehydrogenase, which is involved in synthesis of red eye pigment. If a wild-type copy of the rosy genes is included in the P element vector, it will restore the red eye color. If the offspring of a rosy −/− transgenic fly has red eyes, this implies that the transgene was inserted into the germline, and all the cells in the offspring will have the transgene.