A third mechanism for concentrating CO2 at the site of rubisco is found in crassulacean acid metabolism (CAM). Despite its name, CAM is not restricted to the family Crassulaceae (Crassula, Kalanchoe, Sedum); it is found in numerous angiosperm families. Cacti and euphorbias are CAM plants, as well as pineapple, vanilla, and agave.
Comparison of CAM plants and C-4 Plants
The CAM mechanism is similar in many respects to the C4 cycle.
In C4 plants, formation of the C4 acids in the mesophyll is spatially separated from decarboxylation of the C4 acids and from refixation of the resulting CO2 by the Calvin cycle in the bundle sheath.
In CAM plants, formation of the C4 acids is both temporally and spatially separated.
At night,
CO2 is captured by PEPcarboxylase in the cytosol, and the malate that forms from the oxaloacetate product is stored in the vacuole.
During the day,
the stored malate is transported to the chloroplast and decarboxylated by NADP-malic enzyme, the released CO2 is fixed by the Calvin cycle, and the NADPH is used for converting the decarboxylated triose phosphate product to starch.
The Stomata of CAM Plants Open at Night and Close during the Day
Mechanism:
CAM plants such as cacti achieve their high water use efficiency by opening their stomata during the cool, desert nights and closing them during the hot, dry days. Closing the stomata during the day minimizes water loss, but because H2O and CO2 share the same diffusion pathway, CO2 must then be taken up at night.
- CO2 is incorporated via carboxylation of phosphoenolpyruvate to oxaloacetate, which is then reduced to malate.
- The malate accumulates and is stored in the large vacuoles that are a typical, but not obligatory, anatomic feature of the leaf cells of CAM plants.
- The accumulation of substantial amounts of malic acid, equivalent to the amount of CO2 assimilated at night, has long been recognized as a nocturnal acidification of the leaf.
Day
- With the onset of day, the stomata close, preventing loss of water and further uptake of CO2.
- The leaf cells deacidify as the reserves of vacuolar malic acid are consumed.
- Decarboxylation is usually achieved by the action of NADP-malic enzyme on malate.
- Because the stomata are closed, the internally released CO2 cannot escape from the leaf and instead is fixed and converted to carbohydrate by the Calvin cycle.
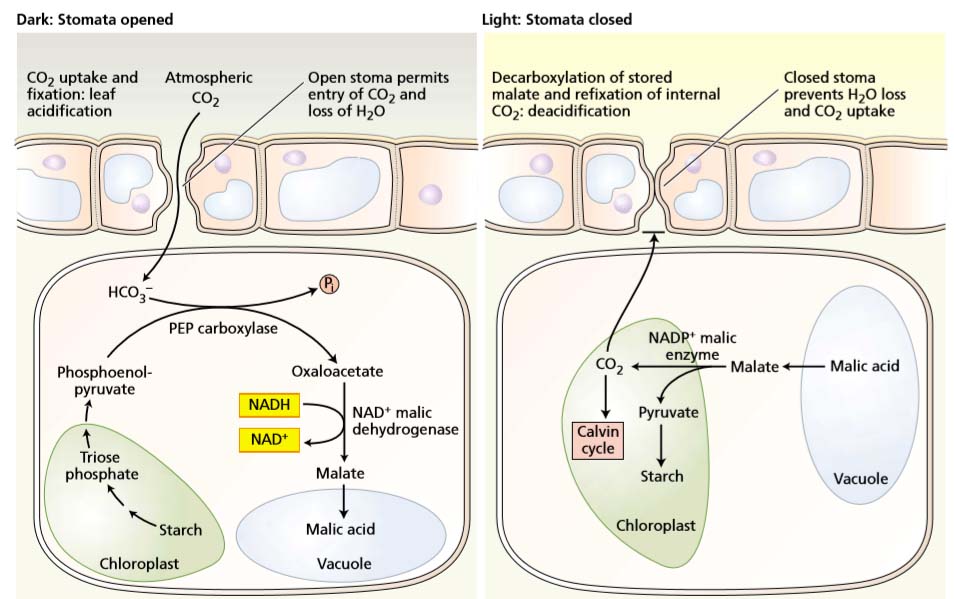
The elevated internal concentration of CO2 effectively suppresses the photorespiratory oxygenation of ribulose bisphosphate and favors carboxylation. The C3 acid resulting from the decarboxylation is thought to be converted first to triose phosphate and then to starch or sucrose, thus regenerating the source of the original carbon acceptor.