Before we see how Watson and Crick solved the structure of DNA, let’s review what was known about genes and DNA at the time that they began their historic collaboration:
1. Genes—
the hereditary “factors” described by Mendel—were known to be associated with specific character traits, but their physical nature was not understood. Similarly, mutations were known to alter gene function, but precisely what a mutation is also was not understood.
2. The one-gene–one-protein theory
postulated that genes control the structure of proteins.
3. Genes were known to be carried on chromosomes.
4. The chromosomes were found to consist of DNA and protein.
5. The results of a series of experiments beginning in the 1920s revealed that DNA was the genetic material.These experiments, described next, showed that bacterial cells that express one phenotype can be transformed into cells that express a different phenotype and that the transforming agent is DNA.
Discovery of transformation
Frederick Griffith made a puzzling observation in the course of experiments on the bacterium Streptococcus pneumoniae performed in 1928.
This bacterium, which causes pneumonia in humans, is normally lethal in mice. However, some strains of this bacterial species have evolved to be less virulent (less able to cause disease or death). In these experiments, Griffith used two strains that are distinguishable by the appearance of their colonies when grown in laboratory cultures.
One strain was a normal virulent type deadly to most laboratory animals. The cells of this strain are enclosed in a polysaccharide capsule, giving colonies a smooth appearance; hence, this strain is identified as S. Griffith’s other strain was a mutant nonvirulent type that grows in mice but is not lethal. In this strain, the polysaccharide coat is absent, giving colonies a rough appearance; this strain is called R. Griffith killed some virulent cells by boiling them.
He then injected the heat-killed cells into mice. The mice survived, showing that the carcasses of the cells do not cause death. However, mice injected with a mixture of heat-killed virulent cells and live nonvirulent cells did die. Furthermore, live cells could be recovered from the dead mice; these cells gave smooth colonies and were virulent on subsequent injection. Somehow, the cell debris of the boiled S cells had converted the live R cells into live S cells. The process, already discussed in Chapter 5, is called transformation.
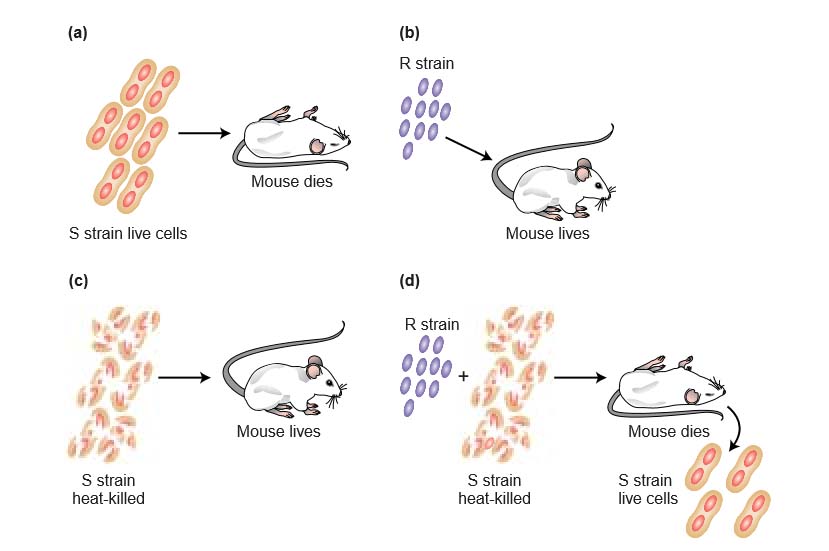
Figure 7-2 The presence of heat-killed S cells transforms live R cells into live S cells. (a) Mouse dies after injection with the virulent S strain. (b) Mouse survives after injection with the R strain. (c) Mouse survives after injection with heat-killed S strain. (d) Mouse dies after injection with a mixture of heat-killed S strain and live R strain. The heat-killed S strain somehow transforms the R strain into virulence.
Transforming Principle
The next step was to determine which chemical component of the dead donor cells had caused this transformation. This substance had changed the genotype of the recipient strain and therefore might be a candidate for the hereditary material.
This problem was solved by experiments conducted in 1944 by Oswald Avery and two colleagues, C. M. MacLeod and M. McCarty . Their approach to the problem was to chemically destroy all the major categories of chemicals in the extract of dead cells one at a time and find out if the extract had lost the ability to transform.The virulent cells had a smooth polysaccharide coat, whereas the nonvirulent cells did not; hence, polysaccharides were an obvious candidate for the transforming agent. However, when polysaccharides were destroyed, the mixture could still transform. Proteins, fats, and ribonucleic acids (RNA) were all similarly shown not to be the transforming agent.
The mixture lost its transforming ability only when the donor mixture was treated with the enzyme deoxyribonuclease (DNase), which breaks up DNA. These results strongly implicate DNA as the genetic material. It is now known that fragments of the transforming DNA that confer virulence enter the bacterial chromosome and replace their counterparts that confer nonvirulence.

Fig: DNA is the agent transforming the R strain into virulence. If the DNA in an extract of heat-killed S strain cells is destroyed, then mice injected with a mixture of the heat-killed cells and the live nonvirulent strain R are no longer killed.
Hershey–Chase experiment
The experiments conducted by Avery and his colleagues were definitive, but many scientists were very reluctant to accept DNA (rather than proteins) as the genetic material. Additional evidence was provided in 1952 by Alfred Hershey and Martha Chase. Their experiment made use of phage T2, a virus that infects bacteria. They reasoned that infecting phage must inject into the bacterium the specific information that dictates the reproduction of new viral particles. If they could find out what material the phage was injecting into the phage host, they would have determined the genetic material of phage.
The phage is relatively simple in molecular constitution. Most of its structure is protein, with DNA contained inside the protein sheath of its “head.”
Hershey and Chase decided to label the DNA and protein by using radioisotopes so that they could track the two materials during infection. Phosphorus is not found in proteins but is an integral part of DNA; conversely, sulfur is present in proteins but never in DNA. Hershey and Chase incorporated the radioisotope of phosphorus (32P) into phage DNA and that of sulfur (35S) into the proteins of a separate phage culture.
Then infected two E. coli cultures with many virus particles per cell: one E. coli culture received phage labeled with 32P and the other received phage labeled with 35S. After allowing sufficient time for infection to take place, they sheared the empty phage carcasses (called ghosts) off the bacterial cells by agitation in a kitchen blender.
They separated the bacterial cells from the phage ghosts in a centrifuge and then measured the radioactivity in the two fractions. When the 32P-labeled phage were used to infect E. coli,most of the radioactivity ended up inside the bacterial cells, indicating that the phage DNA entered the cells.
When the 35S-labeled phages were used, most of the radioactive material ended up in the phage ghosts, indicating that the phage protein never entered the bacterial cell. The conclusion is inescapable: DNA is the hereditary material. The phage proteins are mere structural packaging that is discarded after delivering the viral DNA to the bacterial cell.
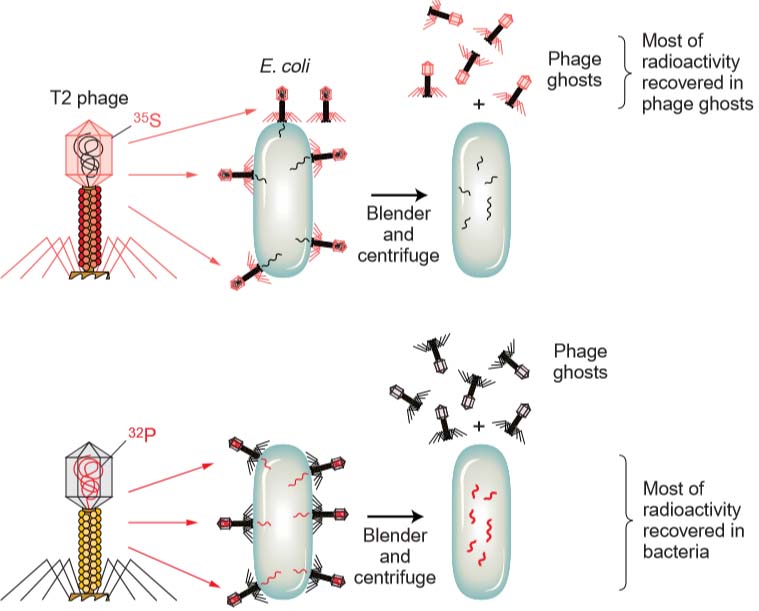
Fig: The Hershey–Chase experiment demonstrated that the genetic material of phage is DNA, not protein. The experiment uses two sets of T2 bacteriophage. In one set, the protein coat is labeled with radioactive sulfur (35S), not found in DNA. In the other set, the DNA is labeled with radioactive phosphorus (32P), not found in protein. Only the 32P is injected into the E. coli, indicating that DNA is the agent necessary for the production of new phage.