Fibers typically are long, spindle-shaped cells, with more or less thick secondary walls, and they usually occur in strands. Such strands constitute the “fibers” of commerce. The process of retting (technical form of the word rotting) used in the extraction of fibers from the plant results in a separation of the fiber bundles from the associated nonfibrous cells. Within a strand, the fibers overlap, a feature that imparts strength to the fiber bundles. In contrast to the thickened primary walls of collenchyma cells, the fiber walls are not highly hydrated. They are therefore harder than collenchyma walls and are elastic rather than plastic. Fibers serve as supporting elements in plant parts that are no longer elongating. The degree of lignification varies, and typically the simple or slightly bordered pits are relatively scarce and slit-like. Many fibers retain their protoplasts at maturity.
Location of fibers in Plant body:
Fibers occur in separate strands or cylinders in the cortex and the phloem, as sheaths or bundle caps associated with the vascular bundles, or in groups or scattered in the xylem and the phloem. In the stems of monocots and dicots the fibers are arranged in several characteristic patterns.
In many Poaceae the fibers form a system having the shape of a ribbed hollow cylinder, with the ribs connected to the epidermis. In Zea, Saccharum, Andropogon, Sorghum , and other related genera the vascular bundles have prominent sheaths of fibers, and the peripheral bundles may be irregularly fused with each other or united by sclerifi ed parenchyma into a sclerenchymatous cylinder. The hypodermal parenchyma may bestrongly sclerified
In some plants no other than the peripheral fibers (primary phloem fibers) occur in the phloem (Alnus, Betula, Linum, Nerium). Others develop fibers in the secondary phloem also, few (Nicotiana, Catalpa, Boehmeria) or many (Clematis, Juglans, Magnolia, Quercus, Robinia, Tilia, Vitis). Some eudicots have complete cylinders of fibers, either close to the vascular tissues (Geranium, Pelargonium, Lonicera, some Saxifragaceae, Caryophyllaceae, Berberidaceae,
Primulaceae) or at a distance from them, but still located to the inside of the innermost layer of the cortex.
Fibers may be prominent in the leaves of monocots. Here they form sheaths enclosing the vascular
bundles, or strands, extending between the epidermis and the vascular bundles, or subepidermal strands not associated with the vascular bundles.
Conifers usually have no fibers in the primary phloem but may have them in the secondary phloem (Sequoia, Taxus, Thuja). Cortical fibers are sometimes present in stems.
Fibers are of Two types: Xylaryand Extraxylary:
Xylary fibers are fibers of the xylem, and extraxylary
fibers are fibers located outside the xylem. Among the
extraxylary fibers are the phloem fibers. Phloem fibers
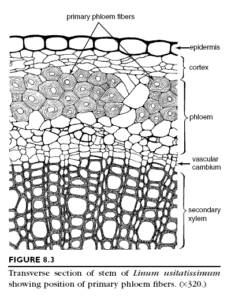
occur in many stems. The flax (Linum usitatissimum)
stem has only one band of fibers, several layers in depth,
located on the outer periphery of the vascular cylinder.These fibers originate in the earliest part of the primary phloem (the protophloem) but mature as fibers after this part of the phloem ceases to function in conduction. Flax fibers are, therefore,
primary phloem fibers, or protophloem fibers. The stems of Sambucus (elderberry), Tilia (basswood), Liriodendron (tulip tree), Vitis (grapevine), Robinia pseudoacacia (black locust), and many others, have both primary phloem fibers and secondary phloem fibers, which are located within the secondary phloem.
Two other groups of extraxylary fibers encountered in the stems of eudicots are the cortical fibers and the perivascular fibers. Cortical fibers, as the name implies, originate in the cortex.
Perivascular fibers are located on the periphery of the vascular cylinder inside the innermost cortical layer. They do not originate as part of the phloem tissue but outside it.
Perivascular fibers are commonly referred to as pericyclic fibers. However, the designation pericyclic is often used with reference to the primary phloem fibers as well.
Extraxylary fibers also include the fibers of the monocots, whether or not associated with the vascular bundles. The cell walls of the extraxylary fibers are often very thick. In the phloem fibers of flax, the secondary wall may amount to 90% of the cross-sectional area of the cell. Not all extraxylary fibers have such wall structure. In mature bamboo culms, some fiber walls show a high
degree of polylamellation, whereas others have no clearly visible lamelae. In addition the secondary walls of the secondary phloem fibers of most woody angiosperms and con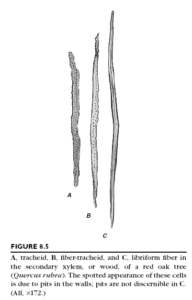 ifers consist of only two layers, a thin outer (S1) layer and a thick inner (S2) layer.
ifers consist of only two layers, a thin outer (S1) layer and a thick inner (S2) layer.
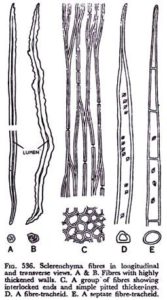
Wood fibers are commonly divided into two main groups, the libriform fi bers and the fiber-tracheids both of which typically have lignified cell walls. The libriform fibers resemble phloem fibers. Libriform is derived from the Latin liber, meaning “inner bark,” that is, phloem. Although the distinction between the two groups of wood fi bers has long been based primarily on the presence of simple pits in libriform fibers and of bordered pits in fi ber-tracheids, truly simple pits in fiber walls are extremely uncommon. The extremes of the two types of wood fi ber are easy to
distinguish, but imperceptible gradations occur between them. Fiber-tracheids also intergrade with tracheids, which have distinctly bordered pits . Commonly the thickness of the wall increases in the sequence of tracheid, fiber-tracheid, and libriform fiber. In addition, in a given sample of wood, the tracheids are usually shorter and the fibers longer, with the libriform fibers attaining the greatest length.
Both Xylary and Extraxylary Fibers May Be Septate or Gelatinous
The phloem and/or xylem fibers of some eudicots undergo regular mitotic divisions after the secondary wall is deposited and are partitioned into two or more compartments by cross-walls, or septae. Such fibers, called septate fi bers, also occur in some monocots where they are nonvascular in origin. The septae consist of a middle lamella and two primary walls and, apparently, they may
or may not be lignified. The septae are in contact but not fused with the secondary wall and are separated by the latter from the original primary wall of the fiber. Apparently the primary walls of the septae continue over part or all of the inner surface of the fiber secondary. Additional secondary wall may develop after the division and cover the septae also In bamboos the septate fibers are characterized by thick polylamellate secondary walls. In addition to a middle lamella and primary wall layers, the septae of these fibers have secondary wall lamellae that continue on the longitudinal
walls of the fibers. Plasmodesmata interconnect the protoplasts via the septae of the fibers, which are living at maturity. Starch commonly is found in septate fibers, indicating a storage function, in addition to a supporting role, for these cells. Some septate fibers also contain crystals of calcium
oxalate.
Both Xylary and Extraxylary Fibers May Be Septate or Gelatinous:
Septate fibers:
The phloem and/or xylem fibers of some eudicots undergo regular mitotic divisions after the secondary wall is deposited and are partitioned into two or more compartments by cross-walls, or septae. Such fibers, called septate fibers, also occur in some monocots where they are nonvascular in origin (in Palmae and Bambuscoideae)
Cell Wall: The septae consist of a middle lamella and two primary walls and, apparently, they may or may not be lignified. The septae are in contact but not fused with the secondary wall and are separated by the latter from the original primary wall of the fiber. Apparently the primary walls of the septae continue over part or all of the inner surface of the fiber secondary wall. Additional secondary wall may develop after the division and cover the septae also. In bamboos the septate fibers are characterized by thick polylamellate secondary walls. In addition to a middle lamella and primary wall layers, the septae of these fibers have secondary wall lamellae that continue on the longitudinal walls of the fibers.Plasmodesmata interconnect the protoplasts via the septae of the fibers, which are living at maturity. Starch commonly is found in septate fibers, indicating a storage function, in addition to a supporting role, for these cells. Some septate fibers also contain crystals of calcium oxalate.
Gelatinous fiber:
Another type of fiber that is neither strictly xylary nor extraxylary is the gelatinous fiber. Gelatinous fibers are identified by the presence of a so-called gelatinousnous layer (G-layer), an innermost secondary wall layer that can be distinguished from the outer secondary wall layer(s) by its high cellulose content and lack of lignin. The cellulose microfi brils of the G-layer are oriented parallel to the long axis of the cell, and hence this layer is isotropic or slightly birefringent when
viewed in transverse section under polarized light. Being hygroscopic, the G-layer has the capacity to absorb much water. Upon swelling, the G-layer may occlude the lumen of the cell; upon drying,
it commonly pulls away from the rest of the wall. Gelatinous fibers have been found in the xylem and phloem of roots, stems, and leaves of eudicots.
Soft and Hard Fibers
The phloem fibers of eudicots represent the bast fibers of commerce. These fibers are classified as soft fibers because, whether or not lignified, they are relatively soft and flexible. Some of the well-known sources and usages of bast fibers are hemp (Cannabis sativa), cordage; jute (Corchorus
capsularis), cordage, coarse textiles; flax (Linum usitatissimum), textiles (e.g., linen), thread; and ramie (Boehmeria nivea), textiles. Phloem fibers of some eudicots are used for making paper.
The fibers of monocots—usually called leaf fibers because they are obtained from leaves—are classified as hard fibers. They have strongly lignified walls and are hard and stiff. Examples of sources and uses of leaf fibers are abaca, or Manila hemp, cordage; bowstring hemp (Sansevieria, entire genus), cordage; henequen and sisal (Agave species), cordage, coarse textiles; New Zealand hemp (Phormium tenax), cordage; and pineapple fiber (Ananas comosus), textiles. Leaf fibers of monocots (together with the xylem) serve as raw material for making paper maize (Zea mays), sugar cane (Saccharum officinarum), esparto grass (Stipa tenacissima), and others.
In commerce, the term fiber is often applied to materials that include, in the botanical sense, other types of cells besides fibers and also to structures that are not fibers at all. In fact the fibers obtained from the leaves of monocots represent vascular bundles together with associated fibers. Cotton fibers are epidermal hairs of seeds of Gossypium raffi a is composed of leaf segments of Raphia palm; rattan is made from stems of Calamus palm.