The two long-distance transport pathways—the phloem and the xylem—extend throughout the plant body. The phloem is generally found on the outer side of both primary and secondary vascular tissues. In plants with secondary growth the phloem constitutes the inner bark.
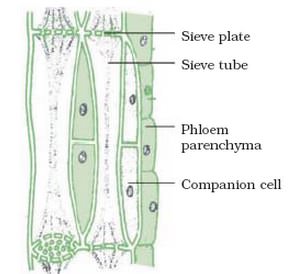
The cells of the phloem that conduct sugars and other organic materials throughout the plant are called sieve elements. Sieve element is a comprehensive term that includes both the highly differentiated sieve tube elements typical of the angiosperms and the relatively unspecialized sieve cells of gymnosperms. In addition to sieve elements, the phloem tissue contains companion cells (discussed below) and parenchyma cells (which store and release food molecules). In some cases the phloem tissue also includes fibers and sclereids (for protection and strengthening of the tissue) and laticifers (latex-containing cells). However, only the sieve elements are directly involved in translocation.
The small veins of leaves and the primary vascular bundles of stems are often surrounded by a bundle sheath, which consists of one or more layers of compactly arranged cells.
Sugar Is Translocated in Phloem Sieve Elements
Early experiments on phloem transport date back to the nineteenth century, indicating the importance of long distance transport in plants. These classical experiments demonstrated that removal of a ring of bark around the trunk of a tree, which removes the phloem, effectively stops sugar transport from the leaves to the roots without altering water transport through the xylem. When radioactive compounds became available, radiolabeled 14CO2 was used to show that sugars made in the photosynthetic process are translocated through the phloem sieve elements.
Many structures normally found in living cells, even the undifferentiated cells from which mature sieve elements are formed. For example, sieve elements lose their nuclei and tonoplasts (vacuolar membrane) during development. Microfilaments, microtubules, Golgi bodies, and ribosomes are also absent from the mature cells. In addition to the plasma membrane, organelles that are retained include somewhat modified mitochondria, plastids, and smooth endoplasmic reticulum. The walls are nonlignified, though they are secondarily thickened in some cases.
Thus the sieve elements have a cellular structure different from that of tracheary elements of the xylem (which are dead at maturity), lack a plasma membrane, and have lignified secondary walls.
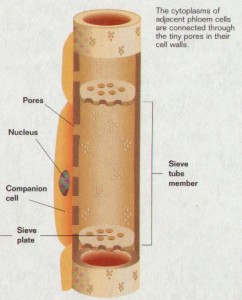
Sieve tube elements found in angiosperms
- Some sieve areas are differentiated into sieve plates; individual sieve tube elements are joined together into a sieve tube.
- Sieve plate pores are open channels.
- P-protein is present in all dicots and many monocots.
- Companion cells are sources of ATP and perhaps other compounds and, in some species, are transfer cells or intermediary cells.
Sieve cells found in gymnosperms
- There are no sieve plates; all sieve areas are similar.
- Pores in sieve areas appear blocked with membranes
- There is no P-protein.
- Albuminous cells sometimes function as companion cells.
Companion Cells
Each sieve tube element is associated with one or more companion cells. The division of a single mother cell forms the sieve tube element and the companion cell. Numerous plasmodesmata penetrate the walls between sieve tube elements and their companion cells, suggesting a close functional relationship and a ready exchange of solutes between the two cells. The plasmodesmata are often complex and branched on the companion cell side.
There are at least three different types of companion cells in the minor veins of mature, exporting leaves: “ordinary” companion cells, transfer cells, and intermediary cells. All three cell types have dense cytoplasm and abundant mitochondria.
Ordinary companion cells have chloroplasts with well-developed thylakoids and a cell wall with a smooth inner surface. Of most significance, relatively few plasmodesmata connect this type of companion cell to any of the surrounding cells except its own sieve element. As a result, the symplast of the sieve element and its companion cell is relatively, if not entirely, symplastically isolated from that of surrounding cells.
Transfer cells are similar to ordinary companion cells, except for the development of fingerlike wall ingrowths, particularly on the cell walls that face away from the sieve element. These wall ingrowths greatly increase the surface area of the plasma membrane, thus increasing the potential for solute transfer across the membrane.
Intermediary cells appear well suited for taking up solutes via cytoplasmic connections . Intermediary cells have numerous plasmodesmata connecting
them to surrounding cells, particularly to the bundle sheath cells. Although the presence of many plasmodesmatal connections to surrounding cells is their most characteristic feature, intermediary cells are also distinctive in having numerous small vacuoles, as well as poorly developed thylakoids and a lack of starch grains in the chloroplasts.