Photoreceptors are defined as pigment molecules that process the energy and informational content of light into a form that can be used by the plant. A pigment that contains protein as an integral part of the molecule is known as a chromoprotein. Thus, photoreceptors typically are chromoproteins. The chromophore (Gr. phoros, bearing) is that portion of the chromoprotein molecule responsible for absorbing light and, hence, colour. The protein portion of a chromoprotein molecule is called the apoprotein. The complete molecule,or holochrome, consists of the chromophore plus the protein.The principal photoreceptors found in plants are described here.
CHLOROPHYLLS ARE PRIMARILY RESPONSIBLE FOR HARVESTING LIGHT ENERGY FOR PHOTOSYNTHESIS
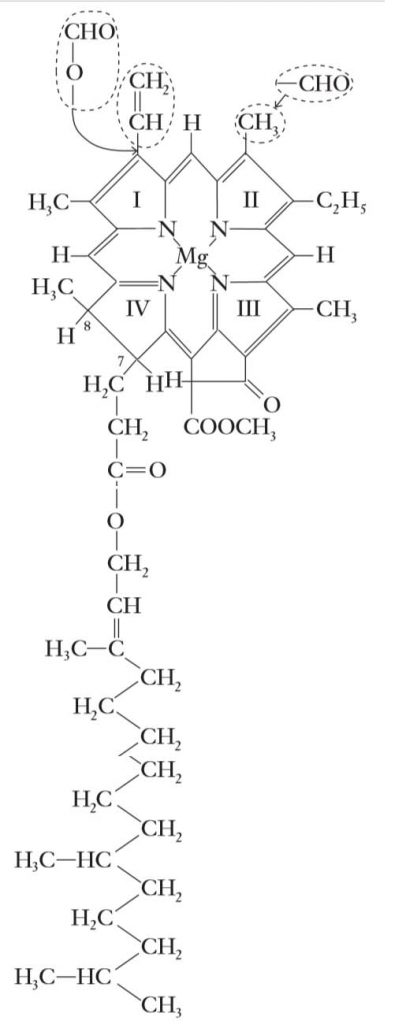
Chlorophyll is the pigment primarily responsible for harvesting light energy used in photosynthesis. The chlorophyll molecule consists of two parts, a porphyrin head and a long hydrocarbon, or phytol tail.
A porphyrin is a cyclic tetrapyrrole, made up of four nitrogen-containing pyrrole rings arranged in a cyclic fashion. Porphyrins are ubiquitous in living organisms and include the heme group found in mammalian hemoglobin and the photosynthetic and respiratory pigments, cytochromes.
Esterified to ring IV of the porphyrin in chlorophyll is a 20-carbon alcohol, phytol. This long, lipid-soluble hydrocarbon tail is a derivative of the 5-carbon isoprene. Isoprene is the precurs or to a variety of important molecules, including other pigments (the carotenes), hormones (the gibberellins), and steroids.
Completing the chlorophyll molecule is a magnesium ion (Mg2+) chelated to the four nitrogen atoms in the center of the ring. Loss of the magnesium ion from chlorophyll results in the formation of a non green product, pheophytin.
Types of chlorophylls
Four species of chlorophyll, designated chlorophyll a, b, c, and d, are known. The chemical structure of chlorophyll a, the primary photosynthetic pigment in all higher plants, algae, and the cyanobacteria, is shown in Figure.
Chlorophyll b is similar except that a formyl group(—CHO) substitutes for the methyl group on ring II. Chlorophyll b is found in virtually all higher plants and green algae,although viable mutants deficient of chlorophyll b are known.
The principal difference between chlorophyll a and chlorophyll c (found in the diatoms, dinoflagellates, and brown algae) is that chlorophyllc lacks the phytol tail.
Finally, chlorophyll d, found only in the red algae, is similar to chlorophyll a except that a (—O—CHO) group replaces the (—CH CH2) group on ring I.
PHYCOBILINS SERVE AS ACCESSORY LIGHT-HARVESTING PIGMENTS IN RED ALGAE AND CYANOBACTERIA
Phycobilins are straight-chain or open-chain tetrapyrrole pigment molecules present in the eukaryotic red algae and the prokaryotic cyanobacteria
Types of Phycobilins
The prefix, phyco, designates pigments of algal origin. Four phycobilins are known. Three of these are involved in photosynthesis and the fourth, phytochromobilin, is an important photoreceptor that regulates various aspects of growth and development.
The three photosynthetic phycobilins are phycoerythrin (also known as phycoerythrobilin), phycocyanin (phycocyanobilin), and allophycocyanin (allophycocyanobilin).
Stucture
In addition to the open-chain tetrapyrrole, the phycobilin pigments differ from chlorophyll in that the tetrapyrrole group is covalently linked with a protein that forms a part of the molecule. In the cell, phycobiliproteins are organized into large macromolecular complexes called phycobilisomes.

The fourth phycobiliprotein, of particular significance to higher plants, is phytochrome, a receptor that plays an important role in many photomorphogenic phenomena.
Interconversion of Phytochrome
Phytochrome (literally, plant pigment) is unique because it exists in two forms that are photoreversible. The form P660 (or Pr) absorbs maximally at 660nm. However, absorption of 660nm light converts the pigment to a second, far-red-absorbing form P735 (or Pfr). Absorption of far-red light by Pfr converts it back to the red-absorbing form. Pfr is believed to be an active form of the pigment that is capable of initiating a wide range of morphogenetic responses.
CAROTENOIDS
Carotenoids comprise a family of orange and yellow pigments present in most photosynthetic organisms.
Occurrence
Found in large quantity in roots of carrot and tomato fruit, carotenoid pigments are also prominent in green leaves. In the fall of the year, the chlorophyll pigments are degraded and the more stable carotenoid pigments account for the brilliant orange and yellow colours so characteristic of autumn foliage.
Chemical nature
Because the carotenoids are predominantly hydrocarbons, they are lipid soluble and found either in the chloroplast membranes or in specialized plastids called chromoplasts.
Types
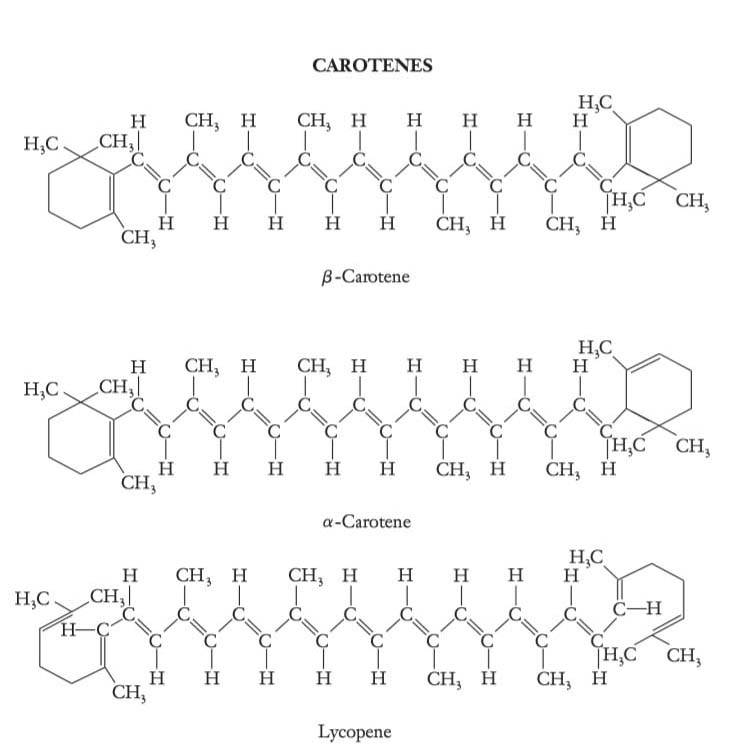
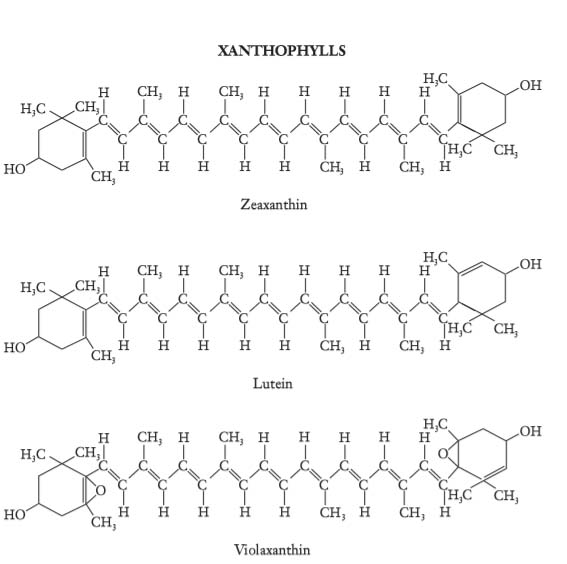
The carotenoid family of pigments includes carotenes and xanthophylls
Carotenes are predominantly orange or red-orange pigments. β-carotene is the major carotenoid in algae and higher plants. Note that in β-carotene and α-carotene (a minor form), both ends of the molecule are cyclized. Other forms, such as γ-carotene, found in the green photosynthetic bacteria, have only one end cyclized. Lycopene, the principal pigment of tomato fruit, has both ends open.
The yellow carotenoids, xanthophylls, are oxygenated carotenes.