Occurence:
Puccinia includes about 700 species which are important as they cause rust dieseases of economically important crop plants such as wheat,barley, eats etc.
The most important of all is P. graminis. It is an internal obligate parsite and is found in associated with wheat growing areas. It is an heteroecious (requiring two hosts; wheat and barbery for the completion of life cycle
Wheat plant is called as Primary host and barberry is the secondary host or alternate host
it is a macrocyclic and polymorphic rust, in which the life cycle involves all five spores with a different function.
Plant Body:
The mycelium of Puccinia is well developed and consists of hyphae, which are spetate and intercellular. It obtains nourishment by small round or branched haustoria. There is a single central dolipore in each septum for the protoplasmic connection.
During the life cycle, two types of mycelia are produced, the monokaryotic, mycelium and dikaryotic mycelium.
The dikaryotic mycelium occurs in primary host (Wheat) and the monokaryotic type is found in alternate host ( barberry)
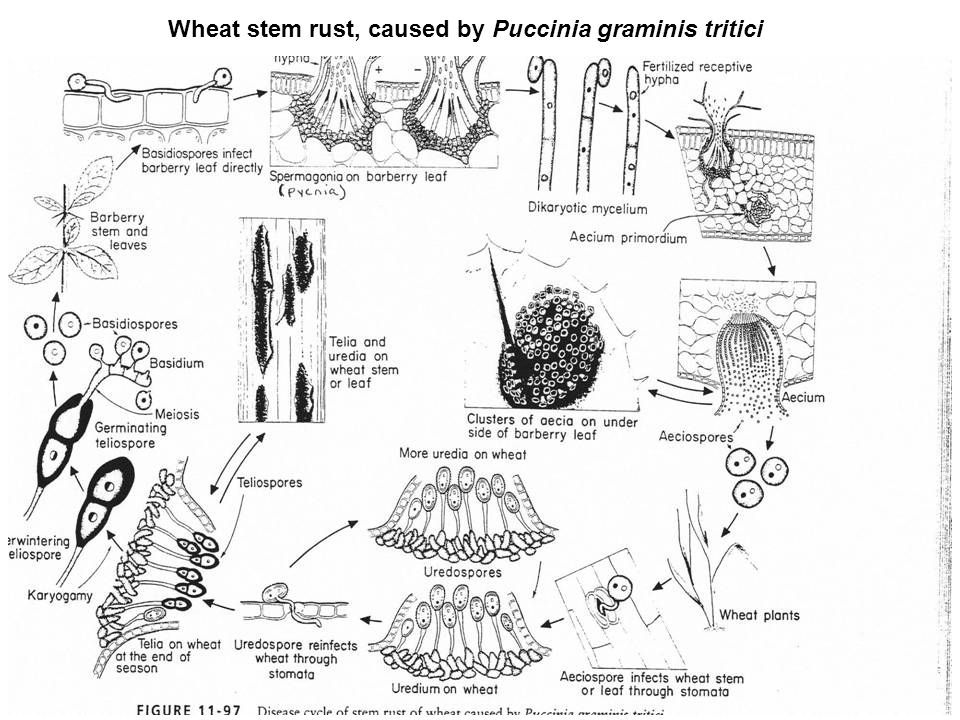
Life Cycle:
Uredospores
Germination of spores
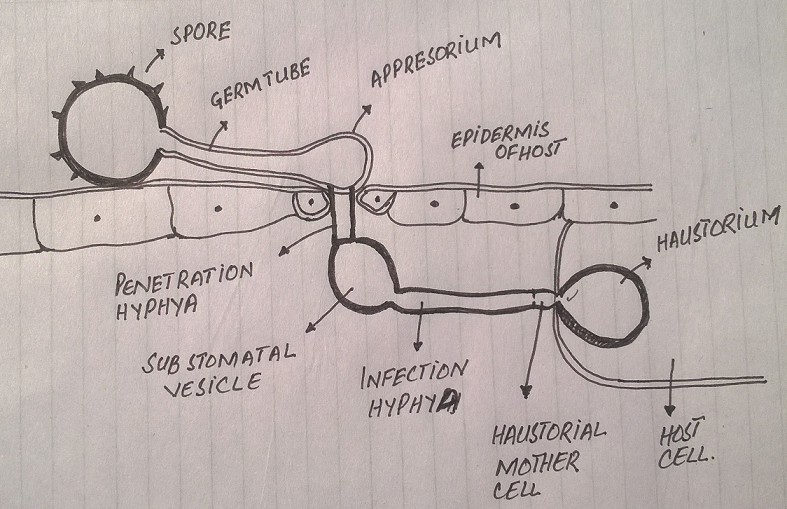
The infection begins in the leaf as the fungus enters through stomata/ injury. The mycelium develops in the host intercellular spaces taking nutrition from host cells through haustoria. Soon (1–2 weeks after infection) they start collecting below the epidermis in clusters called uredosori. Short, erect hyphae called uredinia are produced by the fungal mycelia. The uredinia function as conidiophores and form Urediniospores /uredospores from their tips.
The urediniospores are dikaryotic, oval, stalked; wall is thick spiny/echinulate and brick-red/rust in color.
When a large number of spores form, they exert pressure on the host epidermis and cause its rupture. This exposes the spores and facilitates their dispersal by wind. This appears as rust or brown colored pustules or lesions on the host. The infection first appears on the leaf then goes to the stem, glumes and awn.
Each urediniospore has two germpores (where the wall is thin). The urediniospores germinate by forming a germ tube when it comes in contact with a compatible host. The germ tube produces appressoria which in turn develop the infection peg. The infection pegs enter the host through stomata/injury and finally hyphal strands develop and hyphae spread intercellularly. When fully established, the uredosori are developed again. Urediniospores are the only type of spores which can re-infect the host.
Urediniospores spread from one wheat plant to another through wind, thus spreading the infection from plant to plant, and, field to field. This phase can rapidly spread the infection over a wide area.
Teleutospores:
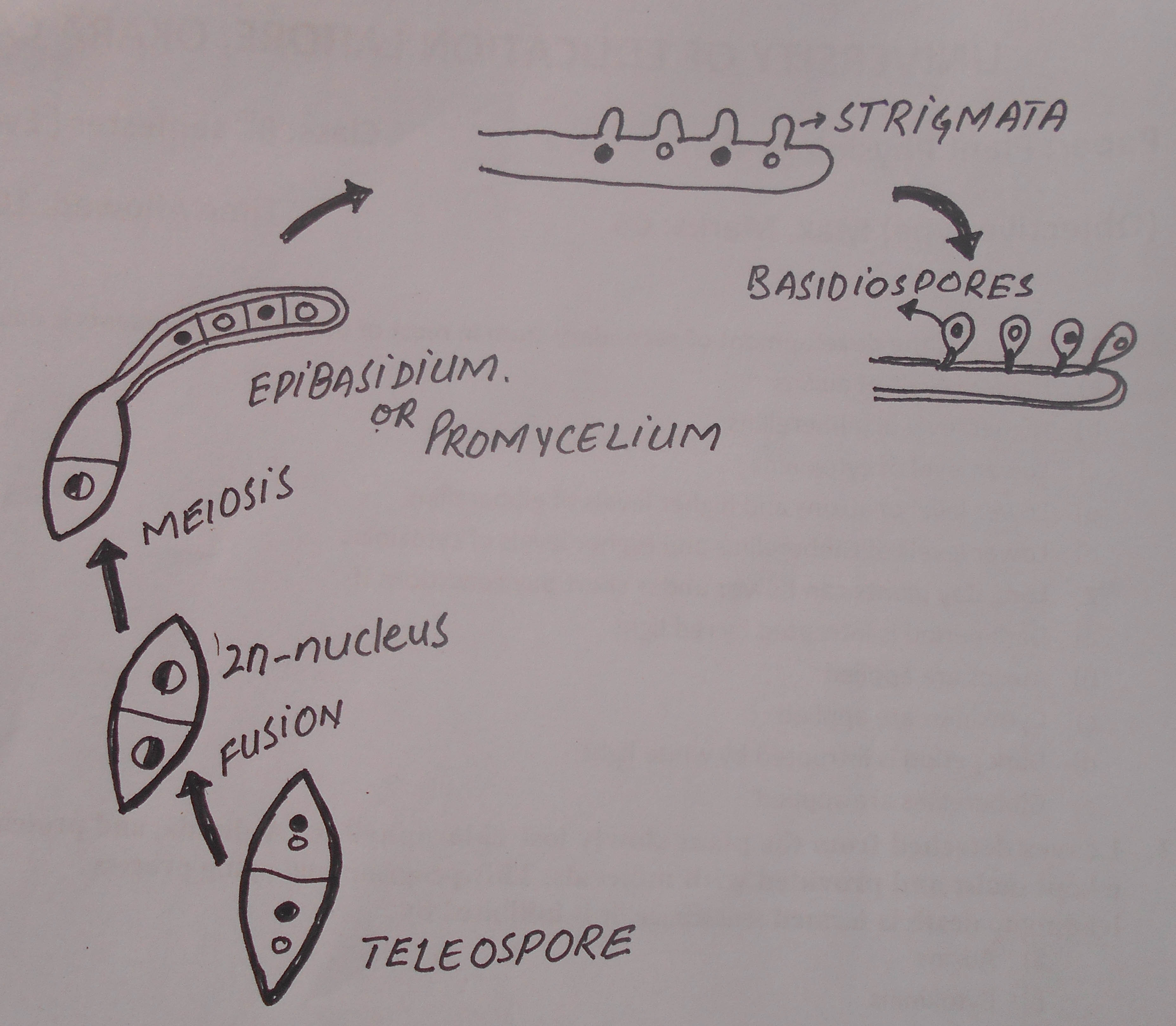
Towards the end of the cereal host’s growing season, the mycelia produce structures called telia. Telia are produced in the same sorus as the uredinia. Telia produce a type of spore called teliospore. These are bicelled, black, thick, smooth walled spores.
They are the only form in which Puccinia graminis is able to overwinter independently of a host. They remain with the straw after harvesting where karyogamy occurs and the teliospores become diploid (2n). The teliospores germinate after a long resting period and exposure to freezing temperature.
Basidiospores:
Germination of teliospores and formation of basidiospores
The upper cell of the spore has an apical germpore, while the lower cell has two laterally apical germpores. A thin hypha comes out of the pore and is called the promycelium. The teliospore is the site of karyogamy and meiosis. Before germination the two nuclei fuse and the resultant diploid (2n) nucleus of the spore undergoes meiosis producing four haploid nuclei. These nuclei migrate into the promycelium, which then becomes septate. This four celled structure is the basidium. It is a septate, uninucleate phragmobasidium .Each cell produces a single haploid basidiospore on sterigmata. Basidiospores are thin-walled and colorless. They cannot infect the cereal host, but can infect the alternative host (Usually barberry). They are usually carried to the alternative host by wind.
Pycniospores:
Once basidiospores arrive on a leaf of the alternative host, they germinate to produce a haploid mycelium which directly penetrates the epidermis and colonizes the leaf. Once inside the leaf the mycelium produces specialized structures called pycnia/spermogonia. The pycnia are flask shaped structures. The pycnia look like small orange bumps on the leaf surface. They produce two types of haploid gametes, the pycniospore/spermogonia and the receptive hyphae. The spermogonia are produced at the tip of short, erect, unbranched hyphae which line the base of the spermogonium. They are formed in large numbers and released from the ostiole in a drop of sticky honeydew which attracts insects. The spermatia function as the male cells. In the neck of the pycnium, long, thin hyphae develop. They grow out of the pycnium through the ostiole and may branch a few times. These are called receptive hyphae. They function as the female gamete.
Insects carry spermatia from one leaf to another; splashing raindrops can also spread spermatia.
Spermatia can fertilize a receptive hypha of the opposite mating type, leading to the production of a dikaryotic mycelium. This is the sexual stage of the life cycle and cross-fertilization provides an important source of genetic recombination.
Aecidiospores:
This dikaryotic mycelium then moves through the leaf tissue and reaches the lower surface, here they forms structures called aecidio mother cells, which produced a type of dikaryotic spores called aecidiospores. These spores have a warty appearance, hexagonal shape, and are formed in chains. The chains of aecidiospores are surrounded by a bell-like structure called aecidial cup. The aecidial cup is emergent i.e. half in and half out of the leaf and is made up of monokaryotic fungal cells. It looks like small orange colored cup like structure on the undersurface of the leaf.
The aecidiospores are able to germinate on the cereal host but not on the alternative host. They are carried by wind to the cereal host where they germinate and the germ tubes penetrate into the plant. The fungus grows inside the plant as a dikaryotic mycelium. Within 1–2 weeks the mycelium produces uredinia and the cycle is complete.
Life cycle of Puccinia