The water content and the rate of water movement in soils depend to a large extent on soil type and soil structure. At one extreme is sand, in which the soil particles may be 1 mm or more in diameter.
Sandy soils have a relatively low surface area per gram of soil and have
large spaces or channels between particles.
At the other extreme is clay, in which particles are smaller than 2 μm in diameter. Clay soils have much greater surface areas and smaller channels between particles. With the aid of organic substances such as humus (decomposing organic matter), clay particles may aggregate into “crumbs” that help improve soil aeration and infiltration of water.

soil through the plant to the atmosphere: differences in
water vapor concentration (Δcwv), hydrostatic pressure
(ΔYp), and water potential (ΔYw).
When a soil is heavily watered by rain or by irrigation, the water percolates downward by gravity through the spaces between soil particles, partly displacing, and in some cases trapping, air in these channels. Water in the soil may exist as a film adhering to the surface of soil particles, or it may fill the entire channel between particles.
In sandy soils, the spaces between particles are so large that water tends to drain from them and remain only on the particle surfaces and at interstices between particles. In clay soils, the channels are small enough that water does not freely drain from them; it is held more tightly.
Field capacity
The moisture-holding capacity of soils is called the field capacity. Field capacity is the water content of a soil after it has been saturated with water and excess water has been allowed to drain away. Clay soils or soils with a
high humus content have a large field capacity. A few days after being saturated, they might retain 40% water by volume. In contrast, sandy soils typically retain 3% water by volume after saturation.
WATER ABSORPTION BY ROOTS
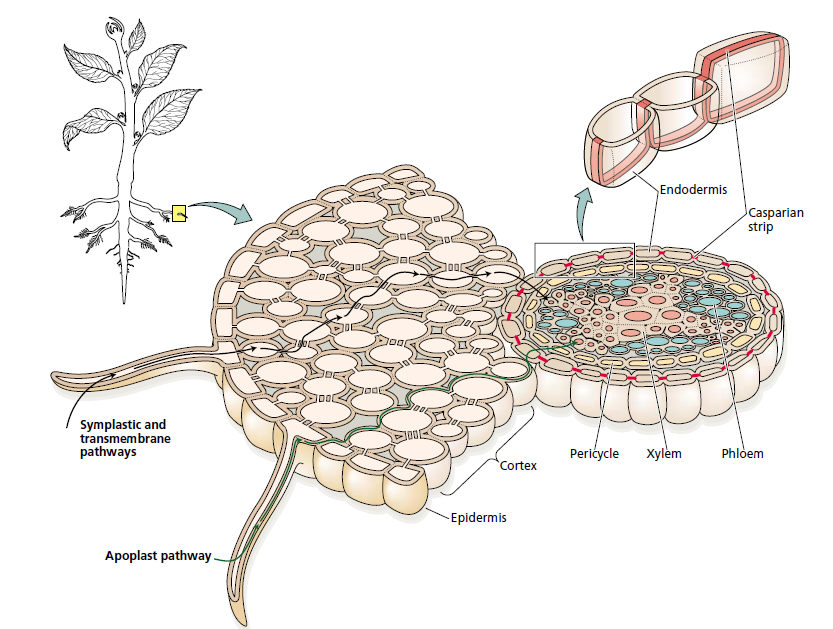
travel via the apoplast pathway, the transmembrane pathway, and the symplast
pathway. In the symplast pathway, water flows between cells through the plasmodesmata
without crossing the plasma membrane. In the transmembrane pathway,
water moves across the plasma membranes, with a short visit to the cell wall space.
At the endodermis, the apoplast pathway is blocked by the Casparian strip.
Intimate contact between the surface of the root and the soil is essential for effective water absorption by the root. This contact provides the surface area needed for water uptake and is maximized by the growth of the root and of root hairs into the soil.
Root hairs are microscopic extensions of root epidermal cells that greatly increase the surface area of the root, thus providing greater capacity for absorption of ions and water from the soil.
Water enters the root most readily in the apical part of the root that includes the root hair zone. More mature regions of the root often have an outer layer of protective tissue, called an exodermis or hypodermis, that contains hydrophobic materials in its walls and is relatively impermeable to water.
Water Pathways in Roots
In the soil, water is transported predominantly by bulk flow. However, when water comes in contact with the root surface, the nature of water transport becomes more complex.
Apoplast Pathway
In the apoplast pathway, water moves exclusively through the cell wall without crossing any membranes. The apoplast is the continuous system of cell walls and intercellular air spaces in plant tissues.
Transmembrane Pathway
The transmembrane pathway is the route followed by water that sequentially enters a cell on one side, exits the cell on the other side, enters the next in the series, and so on. In this pathway, water crosses at least two membranes for each cell in its path (the plasma membrane on entering and on exiting). Transport across the tonoplast may also be involved.
Symplast Pathway
In the symplast pathway, water travels from one cell to the next via the plasmodesmata. The symplast consists of the entire network of cell
cytoplasm interconnected by plasmodesmata.

At the endodermis, water movement through the apoplast pathway is obstructed by the Casparian strip. The Casparian strip is a band of radial cell walls in the endodermis that is impregnated with the waxlike, hydrophobic substance suberin. Suberin acts as a barrier to water and solute movement. The endodermis becomes suberized in the nongrowing part of the root, several millimeters behind the root tip, at about the same time that the first protoxylem elements mature.
The Casparian strip breaks the continuity of the apoplast pathway, and forces water and solutes to cross the endodermis by passing through the plasma membrane. Thus, despite the importance of the apoplast pathway in the root cortex and the stele, water movement across the endodermis
occurs through the symplast.